植物の根毛側面に細胞壁成分を輸送する経路を発見 ~根毛はなぜ細長い形を維持できる?~
植物の根毛は、細長い形状を維持しながら伸長します。これは、先端が伸長すると同時に、根毛側面部分は、二次細胞壁が形成されることで、その膨張が抑制されるためです。しかしながら、根毛側面に二次細胞壁成分を輸送する分子機構については、全く明らかになっていませんでした。この度、金沢大学ナノ生命科学研究所の紺野宏記准教授,中山隆宏准教授,京都府立大学の平野朋子准教授、佐藤雅彦教授らの共同研究グループは、モデル植物シロイヌナズナを用いて、根毛の側面部分に二次細胞壁成分を輸送し、根毛側面を硬くすることで、根毛が細長く真っ直ぐ伸びながら、その形を維持する仕組みを解明しました。
今後、この仕組みを活用して、側面強度を増強した長い根毛を持つ植物体を作出するなど、栄養源が乏しい土壌中から効率よく栄養を吸収できる植物体を開発できる可能性があります。
本研究成果は、国際学術誌 “The Plant Cell”に 2023年9月15日にオンライン掲載されました。
研究概要
植物の根の表皮細胞は、「根毛」を形成する細胞(根毛細胞)と形成しない細胞(非根毛細胞)が交互に配列しています。「根毛」は、根毛細胞の一部が突き出て管状に伸びた構造で、根の表面積を大きくして土壌中の水や養分を吸収する役割があります。
根毛細胞が均等に伸長した場合、風船状に膨らみますが(図1)、実際の根毛は、先端部分が伸びると同時に根毛の側面部分の拡大を抑制するために、細長い管状構造を形成しています。また、土の抵抗に逆らって破けず細長く伸びるために、側面を非常に硬くしています。このような硬い根毛側面には、二次細胞壁が形成されていますが、二次細胞壁成分を輸送する分子機構については、全く不明でした。
今回、京都府立大学を中心とする共同研究グループは、モデル植物シロイヌナズナを用いて、根毛の「先端成長のための物質輸送ルート」と「側面の硬化と成長抑制のための物質輸送ルート」が存在することを発見しました。さらに、これらのルートの担い手は、前者が、SYP132,VAMP721の複合体と、SYP123,VAMP721の複合体で、後者が、SYP123とVAMP727の複合体であることも突き止めました(図2)。
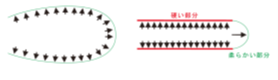
図1.根毛は、内部からの膨圧のはたらきなどにより伸長することから、根毛の細胞壁の硬さが一様な場合、風船状に膨らんでしまい、細長い構造を維持できない。根毛は、側面に二次細胞壁成分が輸送されることで、硬く細長い真っ直ぐな構造が作られる。
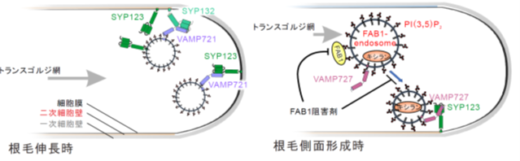
図2. 研究成果の概略図
伸長中の根毛では、SYP123とVAMP721の複合体や、SYP132とVAMP721の複合体が働く「先端成長のための物質輸送ルート」とSYP123とVAMP727の複合体が働く「側面の硬化と成長抑制のための物質輸送ルート」が存在する。
研究の背景
土壌で細長く伸びる根毛は、硬くて、真っ直ぐな構造である。
植物の根毛は、土壌中から水分や養分を効率よく吸収するために発達した細長い構造体で、根の表皮細胞の一部が細長く突出して形成されます。
著者らの先行研究により、根毛の側面形成には、イノシトールリン脂質(注1)の一種であるホスファチジルイノシトール3,5-二リン酸[PI(3,5)P2](注2)とその合成酵素FAB1と低分子量GTPase ROP10が関与することがわかっていました。すなわち、これら分子の複合体形成による「根毛側面の細胞膜直下の表層微小管の構造の安定化」と、 PI(3,5)P2を目印とした「二次細胞壁成分の輸送と分泌」を、PI(3,5)P2が制御し、細長く、真っ直ぐな、側面が硬化した根毛構造が形成されるということです(図3)。しかしながら、PI(3,5)P2が制御する二次細胞壁の輸送と分泌がどのように行われているかについては、ほとんどわかっていませんでした。
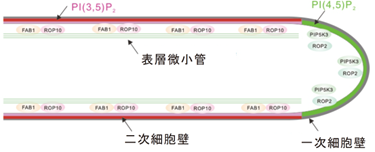
図3.二種類のイノシトールリン脂質のうち、PI(4,5)P2は先端成長に働き、その異性体PI(3,5)P2は根毛側面の硬化に働く。
SYPタンパク質とVAMPタンパク質が物質の輸送場所を決定する。
一方で、著者らは、植物を含む真核生物の細胞内において、膜で囲まれた細胞小器官(オルガネラ)間の物質輸送システムである「小胞輸送」の研究より、SYP132、SYP123というSYPタンパク質とVAMP721、VAMP727というVAMPタンパク質が根毛の伸長に関与することを明らかにしていました。
「小胞輸送」は、「出発地のオルガネラの膜がくびれて、輸送される分子を包んで出芽して輸送小胞が生成され、これが目的地のオルガネラ膜(標的膜)に融合」して、小胞体やゴルジ体で合成された分子を輸送するシステムです。輸送小胞に存在するVAMPタンパク質ファミリーと標的膜に存在するSYPタンパク質ファミリーの組み合わせで、融合オルガネラの特異性、すなわち、物質輸送場所の特異性が決められます(図4)。
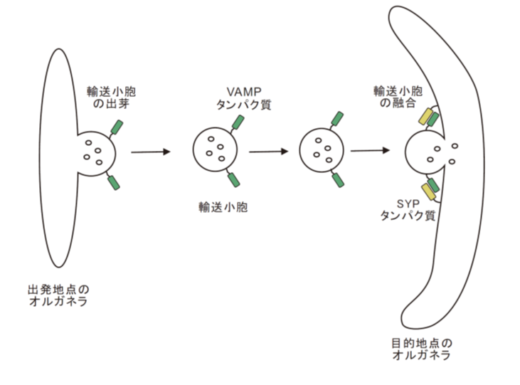
図4.真核生物の細胞内小胞輸送は、「出発地のオルガネラの膜がくびれて、輸送される分子を包んで出芽して輸送小胞が生成され、これが目的地のオルガネラ膜(標的膜)に融合」して輸送するシステム
本研究の成果
SYP123は、FAB1/PI(3,5)P2が制御する輸送経路で働き、二次細胞壁成分を輸送する
根毛の細胞膜に特異的に発現するSYP123と緑色蛍光タンパク質(GFP)を融合したタンパク質 (GFP-SYP123)を発現する形質転換シロイヌナズナの根毛を観察すると、GFP-SYP123は、伸長開始と伸長停止時は、細胞膜全域に均等に配置していましたが、伸長中では、根毛先端よりも側面に強く局在していました(図5)。
そして、GFP-SYP123は、PI(3,5)P2の生成を阻害すると、細胞内に顆粒状に蓄積し、細胞膜に局在できなくなりました。また、SYP123を欠失したシロイヌナズナ(syp123変異体)は、根毛が太くて短い形態を示し、これは野生型でPI(3,5)P2の生成を阻害した場合と同様でした(図6)。
以上のことから、FAB1およびPI(3,5)P2が制御する輸送経路と、SYP123が働く輸送経路は、同一の経路であることが示唆されました。
また、著者らは先行研究において、原子間力顕微鏡(注3)を用いた根毛側面強度の直接的測定法により、PI(3,5)P2の生成を抑制した植物の根毛側面強度が極度に低下していることを示していました。
そこで、syp123変異体の根毛側面強度を測定したところ、やはり著しい低下が観察されました(図7)。これは、syp123変異体の根毛で、二次細胞壁の成分であるキシランとリグニンの蓄積が低下しているためであることが明らかになりました。
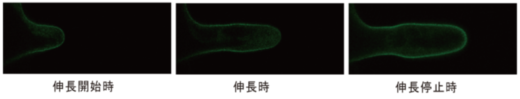
図5.シロイヌナズナ根毛では、GFP-SYP123は、伸長開始時と伸長停止時に細胞膜全体に、伸長時に側面細胞膜に局在する。

図6.PI(3,5)P2合成酵素、FAB1阻害剤YM201636を処理した野生型とsyp123変異体の根毛の形態変化

図7.syp123とvamp727変異体は,根毛側面の硬さが減少している (A)原子間力顕微鏡は、探針を根毛側面に押し付け、カンチレバーのたわみを測定することで、試料の硬さを測定することができる。(B)syp123や vamp727変異体の根毛側面は、野生型の半分の硬さである。
SYP123は、VAP727とペアで二次細胞壁成分を輸送する
一方、二分子蛍光相補 (BiFC)法(注4)と共免疫沈降法(注5)により、SYP123は、輸送小胞に存在するVAMP721とVAMP727の両方と相互作用することを証明しました(図8)。さらに、SYP123とVAMP727の相互作用は、根毛側面の細胞膜上で、二次細胞壁成分を分泌する際に生じることを、共焦点レーザー顕微鏡(注6)と全反射顕微鏡(注7)を用いて示しました(図9)。
これらの結果により、SYP123は、VAMP721が関与する輸送経路とVAMP727が関与する輸送経路の二種類の輸送経路に関与していることが明らかとなりました。
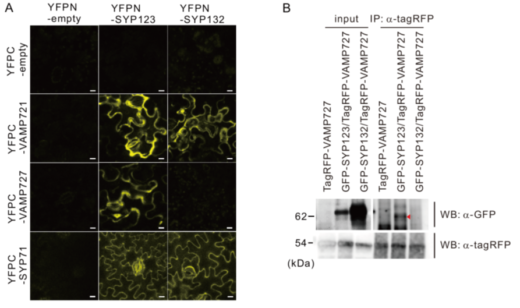
図8.SYP123は、VAMP727とVAMP721の両者と相互作用する (A)二分子蛍光相補(BiFC)法による相互作用解析。SYP123は、VAMP727とVAMP721の両者と相互作用するが、SYP132は、VAMP721のみと相互作用する。(B)共免疫沈降法による相互作用解析。SYP123はVAMP727と相互作用する。
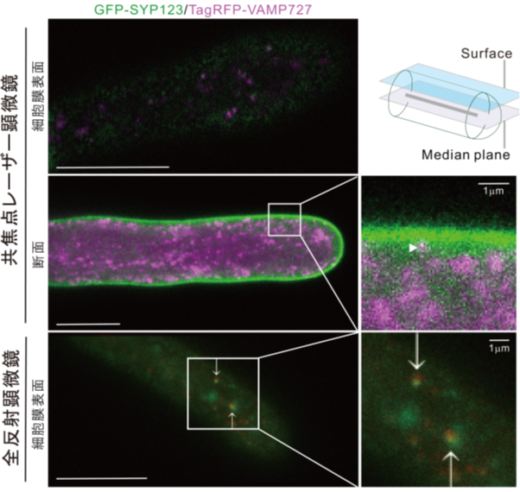
図9.共焦点レーザー顕微鏡と全反射顕微鏡により、GFP-SYP123とTagRFP-VAMP727は、細胞膜周辺で共局在が観察された。
表層微小管は根毛の細長い真っ直ぐな形態をつくる
著者らは以前に、FAB1/PI(3,5)P2の機能阻害が、根毛側面の硬さの低下と細胞膜直下の表層微小管(注8)の断片化を引き起こし、根毛が太くて短く、規則的に波打つ形態を示すことを報告していました。しかし本研究ではsyp123変異体の根毛は、太くて短いものの、波打つ形態は観察されませんでした。そこで、FAB1/PI(3,5)P2の機能阻害植物とsyp123変異体の根毛側面の硬さと表層微小管の安定性をパラメーター化し、力学的な計算機シミュレーションを行い、根毛の形態において、二次細胞壁と表層微小管のそれぞれの寄与を切り分けることに成功しました(図10)。
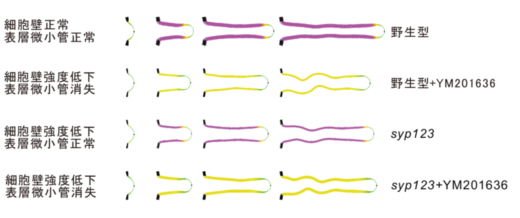
図10.力学的な計算機シミュレーション 根毛では、表層微小管が形態を決め、二次細胞壁の輸送・分泌が形態維持する役割があり、これらの事象は独立している。
以上の結果より、根毛細胞では、「先端成長のための物質輸送ルート」にSYP132とVAMP721の複合体とSYP123とVAMP721の複合体が、「側面の硬化と成長抑制のための物質輸送ルート」に、SYP123とVAMP727の複合体が働くことで、根毛を伸長すると同時に側面を硬くし、根毛の細長い形を作っていることが明らかとなりました。
用語解説
掲載論文情報
- 論文タイトル
- The SYP123-VAMP727 SNARE complex is involved in the delivery of secondary cell wall components for hardening the root hair shank in Arabidopsis
- 著者
- Tomoko Hirano, Kazuo Ebine, Takashi Ueda, Takumi Higaki, Takahiro Watanabe-Nakayama, Hiroki Konno, Hisako Takigawa-Imamura, and Masa H. Sato
- 掲載誌
- The Plant Cell
- 掲載日
- 2023.09.15
- DOI
- 10.1093/plcell/koad240
- URL
- https://doi.org/10.1093/plcell/koad240